
Нотохорд - Notochord
| Нотохорд | |
|---|---|
 Поперечный срез куриного эмбриона инкубации в течение сорока пяти часов.
| |
| Подробности | |
| Предшественник | осевая мезодерма |
| Дает начало | пульпозное ядро |
| Идентификаторы | |
| латинский | нотохорда |
| MeSH | D009672 |
| TE | E5.0.1.1.0.0.8 |
| Анатомическая терминология | |
В анатомии , то хорда представляет собой гибкий стержень выполнен из материала , подобного хрящ . Если у вида есть хорда на любой стадии своего жизненного цикла , это, по определению, хордовая . Хорда лежит вдоль переднезадней оси ( спереди назад ), обычно ближе к дорсальной, чем к вентральной поверхности эмбриона , и состоит из клеток, происходящих из мезодермы .
Наиболее часто упоминаемые функции хорды: как ткань средней линии, которая обеспечивает направляющие сигналы для окружающих тканей во время развития, как место прикрепления мышц и как позвоночный предшественник.
У ланцетников хорда сохраняется на протяжении всей жизни как основная структурная опора тела. У оболочников хорда присутствует только на личиночной стадии, а у взрослого животного полностью отсутствует. Во всех позвоночных , кроме миксин , хорда развивается в позвоночнике , становясь позвонки и межпозвоночные диски центр , который сохраняет структуру , аналогичную исходной хорды.
Состав
Хорда представляет собой длинную палочковидную структуру, которая развивается дорсальнее кишечника и вентральнее нервной трубки. Хорда состоит в основном из ядра гликопротеинов, заключенных в оболочку из коллагеновых волокон, намотанных на две противоположные спирали . Гликопротеины хранятся в вакуолизированных тургидных клетках. Угол между этими волокнами определяет, приведет ли повышенное давление в сердцевине к укорочению и утолщению по сравнению с удлинением и утонением.
Сокращение этих мышечных волокон приводит к движению из стороны в сторону, напоминающему плавание. Усиленная хорда предотвращает движение за счет телескопического движения, как у дождевого червя .
Роль в сигнализации и развитии
Хорда играет ключевую роль в передаче сигналов и координации развития. Эмбрионы современных позвоночных во время гаструляции образуют временные структуры хорды . Хорда находится брюшная в нервную трубку .
Нотогенез - это развитие хорды эпибластами, которые образуют дно полости амниона . Хорд-предшественник происходит из клеток, мигрирующих из примитивного узла и ямки. Хорда формируется во время гаструляции и вскоре после этого вызывает формирование нервной пластинки ( нейруляции ), синхронизируя развитие нервной трубки . На вентральной части нервной борозды происходит осевое утолщение энтодермы . (У двуногих хордовых, например, у людей, эта поверхность правильно называется передней поверхностью). Это утолщение выглядит как борозда (хордальная борозда), края которой анастомозируют (входят в контакт) и таким образом превращают ее в сплошной стержень из клеток многоугольной формы (хорда), который затем отделяется от энтодермы.
У позвоночных, она распространяется по всей длине будущего позвоночного столба, и достигает , как далеко , как передний конец среднего мозга , где она заканчивается в клювообразных конечностях в области будущего Спинка sellae из клиновидной кости . Первоначально он существует между нервной трубкой и энтодермой желточного мешка; вскоре хорда отделяется от них мезодермой , которая растет медиально и окружает ее. Из мезодермы , окружающей нервную трубку и хорды, то череп , позвоночный столб, и мембрана такого мозга и мозгового вещества остистых развиты. Поскольку он происходит от примитивного узла и в конечном итоге располагается в мезодермальном пространстве, считается, что он произошел от мезодермы.
Постэмбриональный остаток хорды обнаруживается в пульпозном ядре межпозвонковых дисков. Изолированные хордальные остатки могут ускользать от их специфического для клонов предназначения в пульпозном ядре и вместо этого прикрепляться к наружным поверхностям тел позвонков , из которых хордальные клетки в значительной степени регрессируют.
У земноводных и рыб
Во время развития земноводных и рыб хорда вызывает развитие гипохорды за счет секреции фактора роста эндотелия сосудов . Гипохорда - это временная структура вентрально от хорды, которая в первую очередь отвечает за правильное развитие дорсальной аорты.
В людях
К 4 годам весь остаток хорды заменяется популяцией хондроцитоподобных клеток неясного происхождения. Сохранение хордовых клеток внутри позвонка может вызвать патологическое состояние: стойкий хордальный канал . Если хорда и носоглотка не разделяются должным образом во время эмбрионального развития, может образоваться депрессия (бурса Торнвальдта) или киста Торнвальдта . Клетки являются вероятными предшественниками редкого рака, называемого хордомой .
Неврология
Исследования хорды сыграли ключевую роль в понимании развития центральной нервной системы . Путем трансплантации и экспрессии второй хорды рядом с дорсальной нервной трубкой, на 180 градусов противоположно нормальному расположению хорды, можно вызвать образование мотонейронов в дорсальной трубке. Формирование двигательных нейронов обычно происходит в брюшной нервной трубке, в то время как дорсальная трубка обычно образует сенсорные клетки .
Хорда секретирует белок, называемый sonic hedgehog (SHH), ключевой морфоген, регулирующий органогенез и играющий важную роль в передаче сигналов о развитии двигательных нейронов. Секреция SHH хордой устанавливает вентральный полюс дорсально-вентральной оси у развивающегося эмбриона.
Эволюция в хордовых

Хорда является определяющим признаком ( синапоморфией ) хордовых и присутствовала на протяжении всей жизни у многих из самых ранних хордовых. Хотя stomochord из полухордовых был когда - то думали, что гомологичные, теперь рассматривается как сходимость . Pikaia, по- видимому, имеет протохорду, и хорды присутствуют в нескольких базальных хордовых, таких как Haikouella , Haikouichthys и Myllokunmingia , все из кембрия .
В ордовикских океанах входят много различных видов бесчелюстной и ранних челюстноротый , которые обладали notochords, либо с прикрепленными элементами костистых или без, прежде всего конодонтами , плакодермами и остракодермы . Даже после эволюции позвоночного столба у chondrichthyes и osteichthyes эти таксоны остались обычными и хорошо представлены в летописи окаменелостей. Некоторые виды (см. Список ниже) вернулись к примитивному состоянию, сохранив хорду во взрослом состоянии, хотя причины этого до конца не изучены.
Сценарии эволюционного происхождения хорды были всесторонне рассмотрены Annona, Holland и D'Aniello (2015). Они указывают на то, что, хотя многие из этих идей не были хорошо поддержаны достижениями молекулярной филогенетики и генетики развития, две из них были фактически возрождены под влиянием современных молекулярных подходов (первый предполагает, что хорда эволюционировала de novo у хордовых. , а второй происходит от гомологичной структуры, аксохорды, которая присутствовала у кольчатых предков хордовых). Выбор между этими двумя сценариями (или, возможно, еще не предложен) должен быть облегчен более тщательными исследованиями сетей регуляции генов у широкого спектра животных.
Постэмбриональная задержка
У большинства позвоночных хорда развивается во вторичные структуры. У других хордовых хорда сохраняется как важная анатомическая структура. Эволюция хорды внутри филума Chordata подробно рассмотрена Холландом и Соморжай (2020).
Следующие организмы сохраняют постэмбриональную хорду:
- Acipenseriformes ( веслонос и осетр )
- Лансолет ( ланцетник )
- Оболочечный (только личиночная стадия)
- Hagfish
- Минога
- Целакант
- Африканская двоякодышащая рыба
- Головастики
- Остракодермы (вымершие)
Внутри Амфиоксуса
Хорда ланцетника выступает за передний конец нервной трубки. Этот выступ служит второй цели, позволяя животному зарываться в отложения на мелководье. Там амфиоксус является фильтрующим питателем и проводит большую часть своей жизни частично погруженным в отложения.
Дополнительные изображения
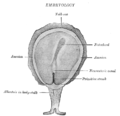
Вид поверхности зародыша гиббона Concolor ( Hylobates concolor ).
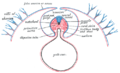
Схема поперечного сечения, показывающая способ образования амниона у цыпленка.

Разрез головы человеческого эмбриона возрастом около двенадцати дней в области заднего мозга.
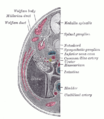
Поперечный разрез человеческого эмбриона от восьми с половиной до девяти недель.




использованная литература
- ^ a b Ствол, Дерек Л. (2005-06-01). «Структура и функция хорды: важный орган для хордовых» . Развитие . 132 (11): 2503–2512. DOI : 10.1242 / dev.01812 . ISSN 0950-1991 . PMID 15890825 .
- ^ Кремер, Jürgen (2009). Заболевания межпозвонкового диска: причины, диагностика, лечение и профилактика . Тиме. С. 15–17. ISBN 978-3-13-582403-1.
- ^ MAR Кель (2000). "Механический дизайн волокнистых гидравлических скелетов: придание жесткости и выпрямление эмбриональных хордов" . Американский зоолог . 40 : 28–041. DOI : 10.1093 / ICB / 40.1.28 .
- ^ a b Хомбергер, Доминик Г. (2004). Рассечение позвоночных . Уокер, Уоррен Ф. (Уоррен Франклин), Уокер, Уоррен Ф. (Уоррен Франклин). (9-е изд.). Бельмонт, Калифорния: Томсон Брукс / Коул. ISBN 0-03-022522-1. OCLC 53074665 .
- ^ «Триламинарный зародышевый диск (3-я неделя)» . www.embryology.ch .
- Перейти ↑ Hood, Rousseaux, Blakley, Ronald D., Colin G., Patricia M. (29 мая 2007 г.). «Эмбрион и плод». Справочник по токсикологической патологии (второе издание) . Academic Press, издательство Elsevier Inc. 2 : 895–936. DOI : 10.1016 / b978-0-12-330215-1.50047-8 . ISBN 9780123302151.CS1 maint: несколько имен: список авторов ( ссылка )
- ^ Генри Грей (1918). Анатомия человеческого тела . Леа и Фебигер. С. 52–54.
- ^ Гэри С. Шенволф; Стивен Б. Блейл; Филип Р. Брауэр; Филиппа Х. Фрэнсис-Уэст (1 декабря 2014 г.). Электронная книга Ларсена по эмбриологии человека . Elsevier Health Sciences. С. 71–72. ISBN 978-1-4557-2791-9.
- ^ Choi, K .; Кон, Мартин Дж .; Харф, Брайан Д. (2009). «Идентификация клеток-предшественников пульпозного ядра и хордовых остатков у мышей: последствия для дегенерации диска и формирования хордомы» . Динамика развития . 237 (12): 3953–3958. DOI : 10.1002 / dvdy.21805 . PMC 2646501 . PMID 19035356 .
- ^ Кливер, Ундина (2000). «Формирование паттерна энтодермы с помощью хорды: развитие гипохорды у Xenopus» (PDF) . Развитие . 127 (4): 869–979. DOI : 10.1242 / dev.127.4.869 . PMID 10648245 .
- ^ Городской, JPG (2000). «Ядро межпозвоночного диска от развития до дегенерации» . Интегративная и сравнительная биология . 40 : 53–061. DOI : 10.1093 / ICB / 40.1.53 .
- ^ Кристоферсон, Lr; Рабин, Б.М.; Hallam, Dk; Рассел, Эй (1 января 1999 г.). «Устойчивость хордального канала: MR и внешний вид простой пленки» (Полный текст) . Американский журнал нейрорадиологии . 20 (1): 33–6. ISSN 0195-6108 . PMID 9974055 .
- ^ Moody MW, Chi DH, Chi DM, Mason JC, Phillips CD, Gross CW и др. (2007). «Киста Торнвальдта: заболеваемость и история болезни» . Ухо Горло Нос J . 86 (1): 45-7, 52. DOI : 10,1177 / 014556130708600117 . PMID 17315835 .
- ^ Пиллаи S, Govender S (2018). «Крестцовая хордома: обзор литературы» . J Orthop . 15 (2): 679–684. DOI : 10.1016 / j.jor.2018.04.001 . PMC 5990241 . PMID 29881220 .
- ^ Echelard, Y; Эпштейн, диджей; Сен-Жак, B; Шен, L; Mohler, J; Mcmahon, Ja; Макмахон, Ап (декабрь 1993 г.). «Sonic hedgehog, член семейства предполагаемых сигнальных молекул, участвует в регуляции полярности ЦНС» . Cell . 75 (7): 1417–30. DOI : 10.1016 / 0092-8674 (93) 90627-3 . PMID 7916661 . S2CID 6732599 .
- ^ Кардонг, Кеннет В. (1995). Позвоночные животные: сравнительная анатомия, функции, эволюция . Макгроу-Хилл. С. 55, 57. ISBN 978-0-697-21991-6.
- ^ "Архивная копия" . Архивировано из оригинала на 2006-03-13 . Проверено 5 сентября 2007 .CS1 maint: заархивированная копия как заголовок ( ссылка )
- ^ "Архивная копия" . Архивировано из оригинала 20 декабря 2010 года . Проверено 21 ноября 2009 года .CS1 maint: заархивированная копия как заголовок ( ссылка )
- ^ Annona, G .; Голландия, Северная Дакота; Д'Аниелло, С. (2015). «Эволюция хорды» . EvoDevo . 6 . Статья 30. DOI : 10,1186 / s13227-015-0025-3 . PMC 4595329 . PMID 26446368 .
- ^ Голландия, Северная Дакота; Соморжай, IML (2020). «Серийный сканирующий электронный микроскоп с блокировкой лица предполагает, что стволовые клетки могут участвовать в росте хорды взрослых хордовых беспозвоночных, багамского ланцетника» . EvoDevo . 11 . Статья 22. DOI : 10,1186 / s13227-020-00167-6 .
- ^ Джозеф Luczkovich; Филип Дж. Мотта; Стивен Ф. Нортон; Карел Ф. Лием (17 апреля 2013 г.). Экоморфология рыб . Springer Science & Business Media. п. 201. ISBN 978-94-017-1356-6.