
Акрокантозавр -Acrocanthosaurus
|
Акрокантозавр |
|
|---|---|
.jpg)
|
|
| Установленный скелет (NCSM 14345) в Музее естественных наук Северной Каролины . | |
|
Научная классификация |
|
| Королевство: | Animalia |
| Тип: | Хордовые |
| Клэйд : | Динозаврия |
| Клэйд : | Заурисхия |
| Клэйд : | Тероподы |
| Семья: | † Carcharodontosauridae |
| Род: |
† Acrocanthosaurus Stovall & Langston , 1950 г. |
| Типовой вид | |
|
† Acrocanthosaurus atokensis Стовалл и Лэнгстон, 1950 г.
|
|
| Синонимы | |
|
|
Acrocanthosaurus ( / ˌ æ к г oʊ ˌ к æ п & thetas ; ə с ɔːr ə с / ак-RO- КАН -thə- SAWR -əs ; означает «высокую-колючки ящерицы») является родом из кархародонтозавриды динозавра , которые существовали вчто сейчас это Северная Америка во время аптского и раннего альбского этапов раннего мелового периода . Как и большинство родов динозавров, Acrocanthosaurus содержит только один вид - A. atokensis . Его окаменелые останки находятся в основном вштатах США Оклахома , Техас и Вайоминг , хотя зубы, приписываемые акрокантозавру , были обнаружены на востоке, вплоть до Мэриленда , что указывает на широкий континентальный ареал.
Акрокантозавр был двуногим хищником . Как следует из названия, он наиболее известен высокими нервными шипами на многих позвонках , которые, скорее всего, поддерживали гребень мышц на шее, спине и бедрах животного. Акрокантозавр был одним из самых крупных теропод, достигавшим 11,5 метров в длину и весом до 6,2 метрических тонны (6,8 коротких тонн). Большие следы теропод, обнаруженные в Техасе, могли быть сделаны акрокантозавром , хотя прямой связи со скелетными останками нет.
Недавние открытия прояснили многие детали его анатомии, что позволило провести специализированные исследования, посвященные структуре мозга и функции передних конечностей. Акрокантозавр был крупнейшим тероподом в своей экосистеме и, вероятно, высшим хищником, который охотился на зауроподов , орнитоподов и анкилозавров .
Открытие и наименование
Acrocanthosaurus назван в честь его высокие остистые отростки, от грек ɑκρɑ / Акра ( 'высокий'), ɑκɑνθɑ / akantha ( 'бельмо' или 'позвоночник') и σɑʊρος / sauros ( 'ящерица'). Существует один названный вид ( A. atokensis ), названный в честь округа Атока в Оклахоме , где были обнаружены оригинальные образцы. Название было придумано в 1950 году американскими палеонтологами Дж. Уиллисом Стоваллом и Ванном Лэнгстоном-младшим. Лэнгстон предложил название «Acracanthus atokaensis» для рода и вида в своей неопубликованной магистерской диссертации 1947 года , но для официальной публикации название было изменено на Acrocanthosaurus atokensis. .

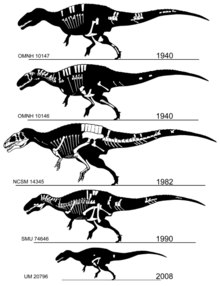
Голотип и паратип ( OMNH 10146 и OMNH 10147), обнаруженный в начале 1940 - х и описана в то же время в 1950 году, состоят из двух частичных скелетов и кусок материала черепа из свиты Antlers в Оклахоме. Два гораздо более полных экземпляра были описаны в 1990-х годах. Первый ( SMU 74646) представляет собой частичный скелет без большей части черепа, извлеченный из формации Твин-Маунтинс в Техасе и в настоящее время являющийся частью коллекции Музея науки и истории Форт-Уэрта . Еще более полный скелет ( NCSM 14345, по прозвищу «Фрэн») был извлечен из формации Рогов в Оклахоме Сефисом Холлом и Сидом Лавом, подготовлен Институтом Блэк-Хиллз в Южной Дакоте и сейчас находится в Музее естествознания Северной Каролины. Наук в Роли . Этот экземпляр является самым крупным и включает единственные известные череп и переднюю конечность полностью. Скелетные элементы OMNH 10147 почти того же размера, что и сопоставимые кости в NCSM 14345, что указывает на животное примерно такого же размера, в то время как голотип и SMU 74646 значительно меньше.
Присутствие Acrocanthosaurus в формации Cloverly было установлено в 2012 году при описании другого частичного скелета, UM 20796. Этот образец, состоящий из частей двух позвонков, частичных лобковых костей , бедренной кости , частичной малоберцовой кости и фрагментов, представляет собой молодой особь. животное. Он пришел из bonebed в Bighorn бассейне северо-центральной части штата Вайоминг , и был найден возле лопатки в виде завропосейдон . Ассортимент других фрагментарных остатков теропод из формации также может принадлежать Acrocanthosaurus , который может быть единственным крупным тероподом в формации Cloverly.
Акрокантозавр может быть известен по менее полным останкам за пределами Оклахомы, Техаса и Вайоминга. Зуб из южной Аризоны был отнесен к этому роду, и соответствующие следы зубов были обнаружены в костях зауроподов из той же области. Несколько зубов из формации Арундел в Мэриленде были описаны как почти идентичные зубам акрокантозавра и могут представлять восточного представителя этого рода. Многие другие зубы и кости из различных геологических образований на западе Соединенных Штатов также были отнесены к акрокантозаврам , но большинство из них были идентифицированы неправильно; Однако есть некоторые разногласия с этой оценкой в отношении окаменелостей формации Cloverly.
Описание

Акрокантозавр был одним из крупнейших известных теропод. Самый крупный из известных образцов (NCSM 14345), по оценкам, имел размеры 11,5 м (38 футов) от носа до кончика хвоста и весил от 5,7 до 6,2 метрических тонны (от 6,3 до 6,8 коротких тонн) с максимальным максимальным весом 7,25 метрических тонн (7,99 тонны). короткие тонны) в пределах возможностей для этого экземпляра. Только его череп был почти 1,3 м в длину.


Череп акрокантозавра , как и большинства других аллозавроидов , был длинным, низким и узким. Отверстие для уменьшения веса перед глазницей ( анторбитальное отверстие ) было довольно большим, более четверти длины черепа и двух третей его высоты. Внешняя поверхность верхней челюсти ( кость верхней челюсти) и верхняя поверхность носовой кости на крыше морды были не такими шероховатыми, как у гиганотозавра или кархародонтозавра . От носовых костей поднимались длинные низкие гребни, идущие вдоль каждой стороны морды от ноздри до глаза, где они продолжались на слезных костях . Это характерная особенность всех аллозавроидов. В отличие от аллозавра , на слезной кости перед глазом не было выступающего гребня. Слезная и заднеглазничная кости соединяются, образуя густую надбровную дугу над глазом, как это видно у кархародонтозаврид и неродственных абелизаврид . Девятнадцать изогнутых зубчатых зубов были выровнены с каждой стороны верхней челюсти, но количество зубов для нижней челюсти не было опубликовано. Acrocanthosaurus зубы были шире , чем у кархародонтозавры и не имеют морщинистую текстуру , которая характеризовала кархародонтозавриду. Зубной кости (зуб подшипника нижней челюсти кость) был квадрат с на переднем крае, как и в Гиганотозавр , и мелкой, в то время как остальная часть челюсти позади него стал очень глубоким. Акрокантозавр и гиганотозавр имели общий толстый горизонтальный гребень на внешней поверхности надугловой кости нижней челюсти под сочленением с черепом.


Самой примечательной особенностью Acrocanthosaurus был ряд высоких нервных шипов, расположенных на позвонках шеи, спины, бедер и верхней части хвоста, которые могли более чем в 2,5 раза превышать высоту позвонков, от которых они отходили. У других динозавров также были высокие шипы на спине, иногда намного выше, чем у акрокантозавров . Например, африканский род Spinosaurus имел шипы высотой почти 2 м (6,6 фута), что примерно в 11 раз выше, чем тела его позвонков. Нижние шипы Acrocanthosaurus имели прикрепления для мощных мышц, как у современных бизонов , вероятно, образуя высокий толстый гребень на его спине. Функция позвоночника остается неизвестной, хотя, возможно, они участвовали в коммуникации , хранении жира , мускулатуре или контроле температуры . Все его шейки (шея) и спинного (назад) позвонки были видные депрессий ( pleurocoels ) по бокам, в то время как каудальной (хвостовой) позвонки буровых мелких. Это больше похоже на кархародонтозавридов, чем на аллозавров .
Помимо позвонков, Acrocanthosaurus имел типичный аллозавроидный скелет. Акрокантозавр был двуногим, с длинным тяжелым хвостом, уравновешивающим голову и тело, а его центр тяжести находился над бедрами. Его передние конечности были относительно короче и крепче, чем у аллозавра, но в остальном были похожи: на каждой руке было по три когтистых пальца. В отличие от многих более мелких быстро бегающих динозавров, его бедренная кость была длиннее большеберцовой кости и плюсневых костей , что свидетельствует о том, что акрокантозавр не был быстрым бегуном. Неудивительно, что кости задних ног Acrocanthosaurus были пропорционально более крепкими, чем его меньший родственник Allosaurus . На каждой ноге было по четыре пальца, хотя, как это типично для теропод, первый был намного меньше остальных и не касался земли.
Классификация и систематика

Acrocanthosaurus это объявление в надсемейство Allosauroidea в инфраотряда тетануры . Это надсемейство характеризуется парными выступами на носовой и слезной костях на вершине морды и высокими нервными шипами на шейных позвонках, среди других особенностей. Первоначально он был помещен в семейство Allosauridae вместе с Allosaurus , эта договоренность также подтверждена исследованиями еще в 2000 году. Большинство исследований показало, что он является членом родственного семейства Carcharodontosauridae.
На момент открытия Acrocanthosaurus и большинство других крупных теропод были известны только по фрагментарным останкам, что привело к весьма разнообразным классификациям для этого рода. J. Willis Stovall и Wann Langston младший первый назначен его на «Antrodemidae», эквивалент аллозавриды , но он был переведен в таксономическую мусорную корзинку Megalosauridae по Альфред Шервуд Ромер в 1956 году других авторов, длинные шипы на его позвонках предложили отношения со спинозавром . Эта интерпретация акрокантозавра как спинозаврида сохранялась до 1980-х годов и повторялась в полутехнических книгах о динозаврах того времени.
Высокие колючие позвонки из раннего мела Англии когда-то считались очень похожими на позвонки Acrocanthosaurus , и в 1988 году Грегори С. Пол назвал их вторым видом этого рода - A. altispinax . Эти кости первоначально были отнесены к Altispinax , английскому тероподу, иначе известному только по зубам, и это назначение привело, по крайней мере, к одному автору, который предположил, что сам Altispinax был синонимом Acrocanthosaurus . Эти позвонки позже были отнесены к новому роду Becklespinax , отдельному от Acrocanthosaurus и Altispinax .


Большинство кладистических анализов , включая Acrocanthosaurus обнаружили , что это кархародонтозавриды, как правило , в базальном положение по отношению к африканским кархародонтозаврам и Гиганотозавру из Южной Америки . Его часто считали сестринским таксоном столь же базальной Eocarcharia , также из Африки. Neovenator , обнаруженный в Англии, часто считается еще более базальным кархародонтозавридом или базальным членом сестринской группы под названием Neovenatoridae . Это говорит о том, что семья возникла в Европе, а затем расселилась по южным континентам (в то время объединилась как суперконтинент Гондвана ). Если бы Acrocanthosaurus был кархародонтозавридом, то расселение также произошло бы в Северной Америке. Все известные кархародонтозавриды жили в период от раннего до среднего мелового периода. Следующая кладограмма Новаса и др. , 2013, показывает размещение Acrocanthosaurus внутри Carcharodontosauridae.
|
|||||||||||||||||||||||||||||||||||||||||||||||||
Палеобиология
Рост и долголетие
Судя по особенностям костей голотипов OMNH 10146 и NCSM 14345, Acrocanthosaurus требовалось не менее 12 лет для полного роста. Это число могло быть намного выше, потому что в процессе ремоделирования костей и роста костномозговой полости некоторые линии Харриса были потеряны. Если учесть эти линии, Acrocanthosaurus потребовалось 18–24 года, чтобы созреть.
Функция передних конечностей

Как и у большинства других нептичьих теропод, передние конечности Acrocanthosaurus не касались земли и не использовались для передвижения; вместо этого они выполняли хищническую функцию. Открытие полной передней конечности (NCSM 14345) позволило провести первый анализ функции и диапазона движений передней конечности у Acrocanthosaurus . В ходе исследования изучались поверхности костей, которые могли бы сочленяться с другими костями, чтобы определить, как далеко суставы могут двигаться без вывиха . Во многих суставах кости не соответствовали друг другу, что указывает на наличие значительного количества хряща в суставах, как это наблюдается у многих современных архозавров . Среди других результатов, исследование показало, что в состоянии покоя передние конечности свисали с плеч, плечевая кость была слегка наклонена назад, локоть согнут, а когти смотрели медиально (внутрь). Плечо акрокантозавра было ограничено в диапазоне движений по сравнению с человеческим. Рука не могла поворачиваться по полному кругу, но могла втягиваться (отклоняться назад) на 109 ° от вертикали, так что плечевая кость могла быть немного наклонена вверх. Вытягивание (качание вперед) ограничивалось только 24 ° относительно вертикали. Рука не смогла достичь вертикального положения , когда аддуктообразовани (качается вниз) , но может похитить (качаться вверх) до 9 ° выше горизонтали. Движение в локтевом суставе также было ограничено по сравнению с людьми: общий диапазон движений составлял всего 57 °. Рука не может полностью продлить (Выпрямление), да и не мог сгибать (изгиб) очень далеко, с плечевой костью не в состоянии даже сформировать правильный угол с предплечьем. Радиус и локтевая кость (кости предплечья) заперли вместе , так что не было никакой возможности пронации или супинаций (скручивание) , как в человеческих предплечьях.
Ни одна из запястных костей (костей запястья) не совмещается точно, что позволяет предположить наличие большого количества хряща в запястье, которое могло бы придать ему жесткость. Все пальцы были способны чрезмерно разгибаться (сгибаться назад), пока почти не касались запястья. При сгибании средняя цифра сходится к первой, а третья - внутрь. На первом пальце руки был самый большой коготь, который был постоянно согнут, так что он загибался назад к нижней стороне руки. Точно так же средний коготь мог быть постоянно согнут, в то время как третий коготь, также самый маленький, мог как сгибаться, так и разгибаться.

После определения диапазона движений в суставах передней конечности, в исследовании была выдвинута гипотеза о хищных привычках Acrocanthosaurus . Передние конечности не могли далеко качнуться вперед, не могли даже поцарапать шею животного. Поэтому маловероятно, что они использовались при первоначальном отлове добычи, а акрокантозавр, вероятно, руководил пастью во время охоты. С другой стороны, передние конечности могли очень сильно втягиваться к телу. Как только добыча была схвачена челюстями, мускулистые передние конечности могли втягиваться, прижимая добычу к телу и препятствуя ее побегу. Когда животное-жертва попыталось отодвинуться, его еще больше накололи на постоянно согнутые когти первых двух пальцев. Чрезвычайная чрезмерная растяжимость пальцев могла быть адаптацией, позволившей Acrocanthosaurus удерживать борющуюся добычу, не опасаясь вывиха. Как только добыча оказалась в ловушке у тела, акрокантозавр, возможно, поразил ее своими челюстями. Другая возможность состоит в том, что акрокантозавр держал добычу в пасти, при этом неоднократно втягивая передние конечности, разрывая большие раны когтями. Другие, менее вероятные теории предполагали, что диапазон движений передних конечностей позволяет хвататься за бок зауроподов и цепляться за них, чтобы опрокинуть зауроподов меньшего роста, хотя это маловероятно из-за того, что акрокантозавр имеет довольно крепкую структуру ног по сравнению с другими аналогично структурированными. теропод.
Строение мозга и внутреннего уха

В 2005 году ученые реконструировали эндокране (реплики) в качестве Acrocanthosaurus полости черепа с помощью компьютерной томографии (КТ) для анализа пространства внутри голотипа (OMNH черепной коробкой 10146). При жизни большая часть этого пространства была бы заполнена мозговыми оболочками и спинномозговой жидкостью , помимо самого мозга. Однако общие характеристики головного мозга и черепных нервов можно определить по эндокасту и сравнить с другими тероподами, для которых были созданы эндокасты. Хотя мозг похож на многие теропод, он больше всего похож на мозг аллозавроидов. Он больше напоминает мозг Carcharodontosaurus и Giganotosaurus , чем мозг Allosaurus или Sinraptor , что подтверждает гипотезу о том, что Acrocanthosaurus был кархародонтозавридом.
Мозг имел слегка сигмовидную форму (S-образную), без значительного расширения полушарий головного мозга , больше напоминал крокодила, чем птицу . Это соответствует общему консерватизму мозга теропод, не относящегося к целурозаврам . У акрокантозавра были большие луковичные обонятельные луковицы , что указывало на хорошее обоняние . Реконструкция полукружных каналов уха, которые контролируют баланс , показывает, что голова держалась под углом 25 ° ниже горизонтали. Это было определено путем ориентации эндокаста так, чтобы боковой полукружный канал был параллелен земле, как это обычно бывает, когда животное находится в настороженной позе.
Возможные следы

В формации Глен Роуз в центральном Техасе сохранилось множество следов динозавров, в том числе большие отпечатки трехпалых теропод. Самая известная из этих троп была обнаружена вдоль реки Пэлюкси в государственном парке Долина динозавров , часть которой сейчас выставлена в Американском музее естественной истории в Нью-Йорке , хотя несколько других мест по всему штату были описаны в литература. Невозможно сказать, какое животное сделало отпечатки, поскольку ископаемые кости не были связаны с тропами. Однако ученые долгое время считали вполне вероятным, что следы принадлежат акрокантозавру . В исследовании 2001 года сравнивали следы Глен Роуз с ногами различных крупных теропод, но не удалось с уверенностью отнести их к какому-либо конкретному роду. Однако исследование отметило, что следы были в пределах ожидаемых для Acrocanthosaurus диапазонов размера и формы . Поскольку формация Глен Роуз близка к формациям Antlers и Twin Mountains как по географическому положению, так и по геологическому возрасту, а единственный крупный теропод, известный из этих формаций, - это Acrocanthosaurus , исследование пришло к выводу, что Acrocanthosaurus, скорее всего , оставил следы.

Знаменитый след Глен Роуз, выставленный в Нью-Йорке, включает в себя следы теропод, принадлежащих нескольким особям, которые двигались в том же направлении, что и двенадцать динозавров- завропод . Отпечатки теропод иногда находят поверх следов зауроподов, что указывает на то, что они были сформированы позже. Это было выдвинуто в качестве доказательства того, что небольшой пакет из Acrocanthosaurus преследовал стадо завроподов. Хотя эта гипотеза интересна и правдоподобна, ее трудно доказать, и существуют другие объяснения. Например, несколько одиночных теропод могли пройти в одном направлении в разное время после того, как зауроподы прошли, создавая впечатление стаи, преследующей свою добычу. То же самое можно сказать и о предполагаемом «стаде» зауроподов, которые также могли или не могли двигаться как группа. В точке, где он пересекает путь одного из зауроподов, на одном из следов тероподов отсутствует след, который был назван доказательством нападения. Однако другие ученые сомневаются в справедливости этой интерпретации, потому что зауропод не изменил походку, как можно было бы ожидать, если бы крупный хищник висел на боку.
Патология
Череп Acrocanthosaurus atokensis голотипом показывает осветить exostotic материал по чешуйчатой . Нейронная позвоночника одиннадцатого позвонка была сломана и зажила в то время как нейронный позвоночник его третьего хвост позвонка был необычным крючок структуры.
Палеоэкология

Определенные окаменелости Acrocanthosaurus были найдены в формации Twin Mountains на севере Техаса, формации Antlers на юге Оклахомы и формации Cloverly в северо-центральном Вайоминге и, возможно, даже в формации Arundel в Мэриленде. Эти геологические образования не были датированы радиометрическим методом , но ученые использовали биостратиграфию, чтобы оценить их возраст. На основании изменений в аммоните таксонов , граница между аптом и альбом этапами в раннем меле была расположена в свите Глена Роуз Техас, который может содержать Acrocanthosaurus следов и лежит чуть выше свиты спаренные гор. Это указывает на то, что формация Twin Mountains целиком лежит в пределах аптского этапа, который длился от 125 до 112 миллионов лет назад. Формация Antlers содержит окаменелости Deinonychus и Tenontosaurus , двух родов динозавров, также обнаруженных в формации Cloverly, которая была радиометрически датирована аптским и альбским этапами, что позволяет предположить схожий возраст Antlers. Следовательно, Acrocanthosaurus, скорее всего, существовал между 125 и 100 миллионами лет назад.
В то время территория, сохранившаяся в формациях Твин-Маунтинс и Рога, была большой поймой , впадающей в мелководное внутреннее море . Через несколько миллионов лет это море расширится на север, превратившись в Западный внутренний морской путь и разделив Северную Америку надвое почти на весь поздний меловой период . Формация Глен Роуз представляет собой прибрежную среду с возможными следами акрокантозавра, сохранившимися в илистых отмелях вдоль древней береговой линии. Поскольку Acrocanthosaurus был крупным хищником, ожидается, что он имел обширный домашний ареал и жил во многих различных средах в этом районе. Потенциальные жертвы включают зауроподов, таких как Astrodon или, возможно, даже огромного Sauroposeidon , а также крупных орнитоподов, таких как Tenontosaurus . Меньший теропод Deinonychus также бродил по местности, но его длина составляла 3 м (10 футов), что, скорее всего, составляло лишь минимальную конкуренцию или даже пищу для Acrocanthosaurus .
использованная литература
внешние ссылки
- Часто задаваемые вопросы об акрокантозаврах из Музея естественных наук Северной Каролины.
- Запись Acrocanthosaurus в базе данных Theropod.
- История « Фран » (NCSM 14345)
- Тропа Глен Роуз в Американском музее естественной истории .